


Az ELKH Agrártudományi Kutatóközpont (ATK) Mezőgazdasági Intézetének munkatársai sikeresen azonosítottak a gabonafélék ivarsejtjeit kialakító számfelező sejtosztódás, azaz a meiózis során a kromatinban végbemenő szerkezetváltozásokat. Az intézetben működő Genetikai Variabilitás Kutatócsoport célja olyan módszerek kidolgozása, amelyek utat nyitnak a korábban nem elérhető, az új környezeti kihívásoknak is megfelelő genetikai kombinációk előállítására a gabonafélék nemesítése során.
Lenykó-Thegze Andrea PhD-hallgató, az ATK tudományos segédmunkatársa, Sepsi Adél és Cseh András ‒ a kutatóközpont tudományos főmunkatársai ‒ témavezetésével a The Plant Journal folyóiratban publikálta legújabb kutatási eredményeit, melyek az apai és anyai kromoszómák gabonafélékben is lejátszódó rekombinációjának részleteit tárják fel. Ezek várhatóan közvetlen hatással lesznek a nemesítés sikerességére – számolt be a magyar sikerről az ELKH weboldala.
A gabonafélék ivarsejtjeit kialakító számfelező sejtosztódás során a kromatinban végbemenő szerkezetváltozásokat azonosítottak az ATK kutatói. Ez azért kiemelkedően fontos, mert a nemesítési programok hatékonysága a kölcsönös genetikai információcsere gyakoriságán és kromoszómán belüli pozícióján alapul. A szülői allélok rekombinációja határozza meg a nemesítés első lépésének a sikerességét. Így döntő szerepe van a termésbiztonság javításában a jelenleg zajló éghajlati változás körülményei között.
Búza és árpa átkereszteződések
A nagyméretű genommal rendelkező gabonaféléinkben, mint például a búzában (Triticum aestivum) és az árpában (Hordeum vulgare) a rekombinációt eredményező kromoszomális átkereszteződések (crossoverek, CO) kizárólag a kromoszómavégekhez közeli szűk régiókban alakulnak ki. A crossoverek kialakulásának e szigorú térbeli szabályozottsága megakadályozza új allélkombinációk kialakulását a kromoszómakarok központi részén és a centromérához közel eső genomi régiókban.
Ezen crossovermentes régiók a genom jelentős hányadát képviselik, és megabázispáronként (Mbp) átlagosan 3‒7 gént tartalmaznak (Lou et al., 2017). Annak érdekében, hogy a homológ rekombináció frekvenciáját és elhelyezkedését befolyásolni lehessen, kulcsfontosságú azon faktorok megértése, amelyek a crossoverek kialakulását szabályozzák.
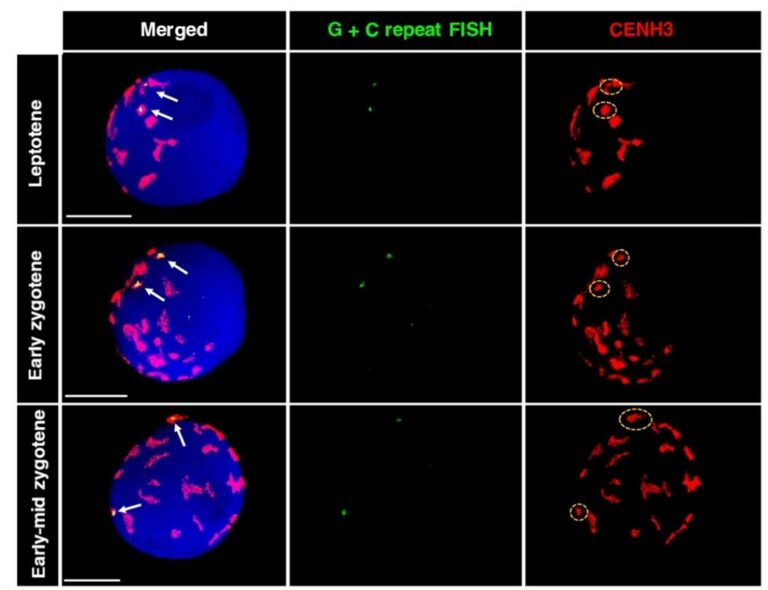
A kutatás két legfontosabb új eredménye a búza genetikai háttérben stabilan öröklődő egyetlen árpakromoszóma-kar dinamikájának feltérképezése során született. Az ImmunoFISH technika alkalmazásával bebizonyosodott, hogy a centromérák a meiózis kezdeti szakaszaiban véletlenszerűen alkotnak csoportokat. Nem történik meg a homológ, illetve homeológ párok egy csoportba rendeződése (1. kép).
Ezt az eltérést fedezték fel a magyar kutatók
A meiotikus rekombináció és a szinapszis folyamatainak párhuzamos nyomon követésével kimutatták, hogy a szubtelomerikus kromatin a rekombináció beindulásakor, a DNS kettős szálú töréseinek kialakulásával egy időben jelentősen meghosszabbodik, és nyitott szerkezetet vesz fel. Ezzel szemben a kromoszómakar és a centromérához közel eső régiók megőrzik zárt kromatinszerveződésüket, és csak később csomagolódnak ki, egyidejűleg a szinaptonémás komplex kezdeti felépülésével és a kettős szálú törések hibajavító folyamatainak beindulásával (2. kép). A zárt pericentromerikus kromatinszerkezet ezeken a kromoszómarégiókon belül hozzájárulhat a crossoverek gátláshoz. Így ennek feloldásához és a rekombináció új genomi régiók felé történő kiterjesztéséhez szükség van az e folyamatokat irányító gének azonosítására.
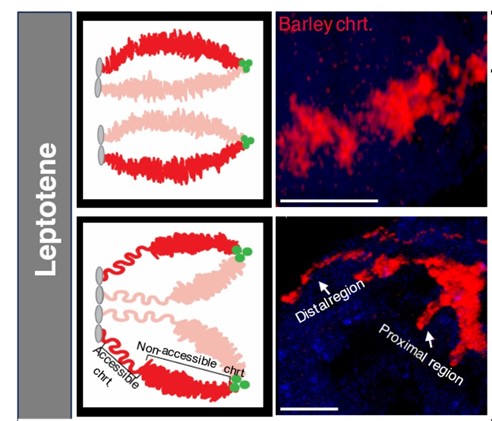
Az 1. és 2. kép forrása: ELKH
Forrás: magro.hu




